
秦巴山区阔叶林地上生物量格局分析
|
姚永慧(1975-),女,湖北安陆人,博士,副研究员,研究方向为地理时空数据分析。 E-mail: yaoyh@lreis.ac.cn |
收稿日期: 2023-12-14
录用日期: 2024-06-05
网络出版日期: 2024-11-07
基金资助
国家自然科学基金项目(41871350)
科技基础资源调查专项(2017FY100900)
Analysis of broadleaved forest aboveground biomass in the Qinling-Daba Mountains
Received date: 2023-12-14
Accepted date: 2024-06-05
Online published: 2024-11-07
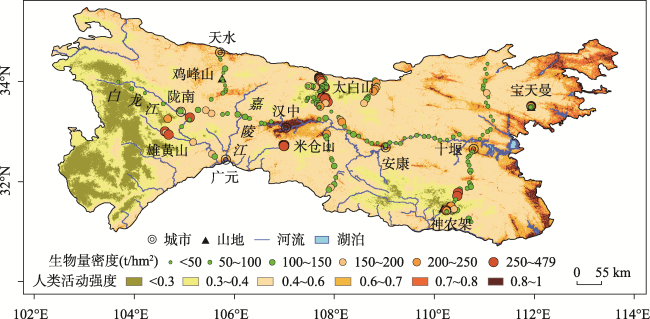
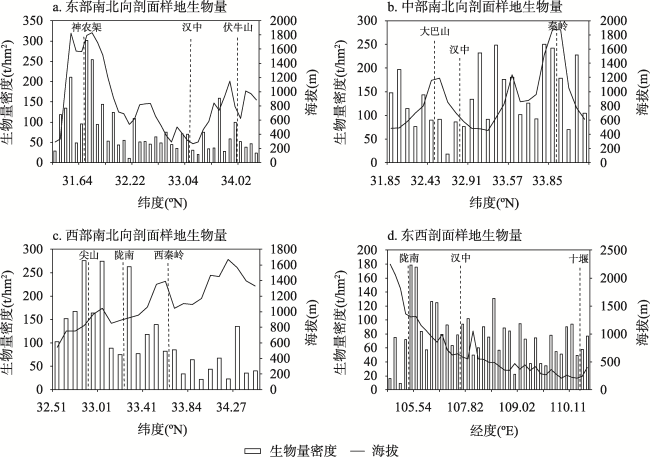
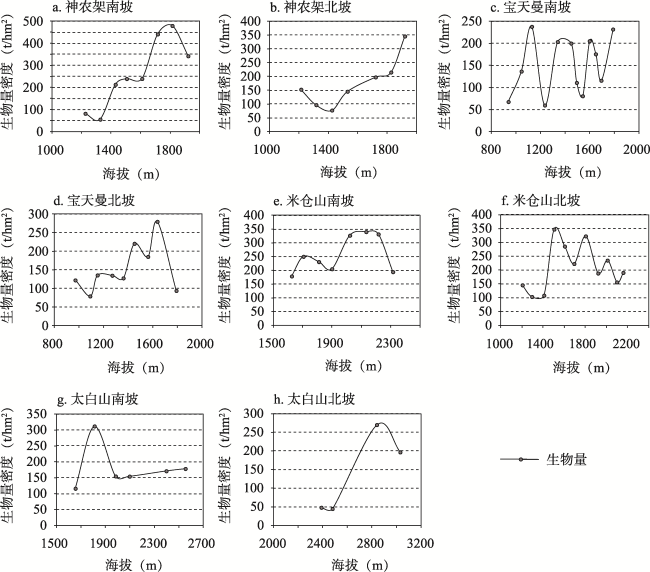
秦巴山区是我国重要的地理-生态过渡带,发育了多样而独特的森林生态系统,其森林生态系统及固碳潜力等关系到区域生态平衡及国家双碳政策的具体布局。阔叶林是森林的主要类型之一,目前秦巴山区森林地上生物量研究集中于太白山、神农架、火地塘林区、秦岭中段等地,或只关注单一或几种典型树种的生物量,尚未从区域上系统分析和估算阔叶林生物量的状况。因此,本文利用“中国南北过渡带综合科学考察”获取的267个阔叶林样方数据,基于统计分析方法研究阔叶林地上生物量的空间分布格局;在此基础上利用相关分析及地理探测器法研究地形、气温、降水及人类活动强度(HAI)4个因素对阔叶林地上生物量的影响。研究结果表明: ① 秦巴山区东部阔叶林地上生物量低,中、西部高;经度方向上呈两种变化模式即南、北两侧山系的生物量自西向东逐渐升高,中部(陇南-汉中-十堰)生物量自西向东逐渐降低;纬度方向上自南向北呈“上升-下降-上升-下降”的变化模式。垂直方向上,阔叶林地上生物量表现为随海拔升高呈“先上升-后下降”的变化模式。② 区域内阔叶林地上生物量格局是人类活动与自然因素相互作用的结果,各因子的影响作用表现为气温>地形>HAI>降水,即除气温、降水和地形等对植物生长发育、分布起重要作用的因素外,人类活动对区域内阔叶林地上生物量的影响不容忽视。本研究可以为秦巴山区的森林生态系统及其碳存储能力、气候变化等研究提供支撑,也能为国家及区域生态规划、环境-经济协调发展提供科学依据。
姚永慧 . 秦巴山区阔叶林地上生物量格局分析[J]. 地理研究, 2024 , 43(11) : 3045 -3057 . DOI: 10.11821/dlyj020231135
The Qinling-Daba Mountains are an important geographic-ecological transitional zone in China, developing diverse and unique forest ecosystems, whose forest ecosystems and carbon sequestration potentials are related to the regional ecological balance and the specific layout of the national dual-carbon policy. At present, the studies on forest aboveground biomass in this area have mainly focused on the local areas, such as Mt. Taibai, Mt. Shennongjia, Huoditang forest area, and the central part of the Qinling Mountains, or only focused on the biomass of a single or a few species, which resulted in fragmented results and could not comprehensively and accurately reflect the overall situation of forest aboveground biomass (AGB) in this area. Therefore, this study utilized the 267 broadleaved forest samples obtained from the "Comprehensive Scientific Investigation of China's North-South Transitional Zone" to investigate the spatial distribution of broadleaved AGB and its environmental influencing factors. The results showed that: (1) Broadleaved forest AGB was low in the eastern Qinling-Daba Mountains and high in the central and western parts; two longitudinal distribution patterns were discovered for broadleaved forest AGB in this area, and one was that the biomass in the mountains located in the north and south of this area gradually increased from west to east, and the other was that the biomass in the central part (Longnan-Hanzhong-Shiyan) gradually decreased from west to east; and the latitudinal distribution pattern of broadleaved forest AGB showed a trend of "increasing-decreasing-increasing-decreasing" from south to north. The altitudinal distribution pattern showed a "first increasing-then decreasing" pattern with increasing altitude. (2) The pattern of broadleaved forest AGB was the result of the interaction between human activities and natural factors, and the influence of each environmental factor was manifested as temperature > terrain > HAI > precipitation, in addition to the factors that play an important role in the growth, development and distribution of plant, such as temperature, precipitation, and terrain, the influence of human activities (HAI) on the broadleaved forest AGB in the study area should be paid more attention. The results can provide support for research on forest ecosystems and their carbon storage capacity, climate change in the Qinling-Daba Mountains, and also provide scientific basis for national and regional ecological planning and environment-economy coordinated development.
表1 基于地面实测法的秦巴山区森林生物量相关研究Tab. 1 Studies on forest biomass in Qinling-Daba Mountains based on ground-survey method |
| 文献(时间) | 森林类型 | 研究地点 | 研究结果与结论 |
|---|---|---|---|
| 肖瑜[20](1989年) | 油松人工林 | 陕西秦岭 | 秦岭油松人工林的生物量较其它省份高 |
| 李晶晶等[21] (2013年) | 华山松林 | 秦岭中段南坡 | 秦岭中段南坡华山松林总生物量81.39 t/hm2,其中乔木层生物量79.45 t/hm2;乔木层在华山松天然林生物量占主要地位 |
| 傅志军[22] (1994年) | 太白红杉林 | 太白山南北坡 | 太白红杉的总生物量为205.55 t/hm2,其中乔木层166.57 t/ hm2 |
| 傅志军[23] (1994年) | 红桦林 | 太白山南北坡 | 红桦林的总生物量为286.79 t/ hm2,其中乔木层251.134 t/ hm2 |
| 李亮等[24] (2013年) | 太白红杉、 巴山冷杉 | 太白山南北坡 | 太白红杉南北坡生物量的平均值为2.91 kgC/m2,巴山冷杉为2.02 kgC/m2,太白红杉生物量的年际波动较大,巴山冷杉生物量的年际波动相对较小,北坡生物量水平大于南坡 |
| 杨凤萍等[25] (2014年) | 油松、华山松 | 秦岭火 地塘林区 | 不同海拔油松和华山松林乔木层生物量和蓄积量增长迅速,且油松的增速一直快于华山松:油松林生物量从1977年的44.40 t/hm2增长到2011年的214.67 t/hm2,华山松林的生物量从1977年的16.74 t/hm2增长到2011年的182.75 t/hm2。环境差异、人为干扰是造成不同海拔间林分生物量、蓄积量产生差异的主要因素;生物学特性和林分环境的共同作用导致了不同森林类型生物量和蓄积量的差异 |
| 陈存根[26] (1996年) | 锐齿槲栎林 | 秦岭南坡 | 锐齿栎林的平均蓄积量为181.2 m3/hm2,平均现存量为208.4 t/hm2,叶面积指数5.631,平均生产量17.97t/hm2,秦岭锐齿栎林具有较高的生物量 |
| 程堂仁等[27] (2007年) | 锐齿槲栎、油松、栓皮栎、杨桦、落叶松、华山松、云冷、其他到时候阔混交林 | 秦岭西段 甘肃小陇山 | 小陇山林区锐齿栎、油松、栓皮栎、杨桦、落叶松、华山松、云冷杉、其他硬阔混交林等8类林分生物量依次为: 84.0472 t、62.4424 t、81.7747 t、77.4367 t、68.9982 t、70.0695 t、96.4865 t、98.7235 t/hm2;各树种的单木生物量、乔木层生物量与胸径和树高之间均存在着紧密的相关关系 |
| 王向雨等[28] (2007年) | 米心水青冈林、锐齿槲栎林 | 神农架地区 | 米心水青冈林的生物量在(251.30~358.63)t/hm2之间,平均为288.70 t/hm2;锐齿槲栎林群落生物量在(134.85~301.20)t/hm2,平均为231.60 t/hm2;两种类型森林生物量的95%以上集中在乔木层 |
| 尉文[29] (2020年) | 锐齿槲栎 | 太白山 | 锐齿槲栎原始林和次生林的平均地上生物量分别为279.50 t/hm2、217.1 t/hm2,生境对生物量具有较大的影响 |
| 张九东[30] (2014年) | 红豆杉 | 陕西省 秦巴山地 | 秦岭北坡的平均生物量为1.096 t/hm2,秦岭南坡的平均生物量为3.91 t/hm2;大巴山北坡的平均生物量为1.38 t/ hm2 |
| 李鹏[31] (2013年) | 森林地上生物量 | 秦岭南坡 | 森林样地平均生物量在(36.78~114.15)t/hm2之间,平均值为73.97 t/hm2;地形因子与生物量具有显著的相关性,随海拔和坡度有先升高后减小的趋势 |
表2 阔叶林分省混合种(组)生物量方程Tab. 2 Mixed species (group) biomass equation of broadleaved forest by province |
| 省份 | 生物量方程 | 拟合R2 | 胸径范围(cm) |
|---|---|---|---|
| 陕西 | 0.96 | 5~70 | |
| 0.74 | |||
| 0.87 | |||
| 河南 | 0.96 | 5~70 | |
| 0.74 | |||
| 0.87 | |||
| 湖北 | 0.98 | 5~81 | |
| 0.94 | |||
| 0.71 | |||
| 四川 | 0.92 | 5~56 | |
| 0.90 | |||
| 0.96 | |||
| 甘肃 | 0.99 | 5~80 | |
| 0.89 | |||
| 0.67 |
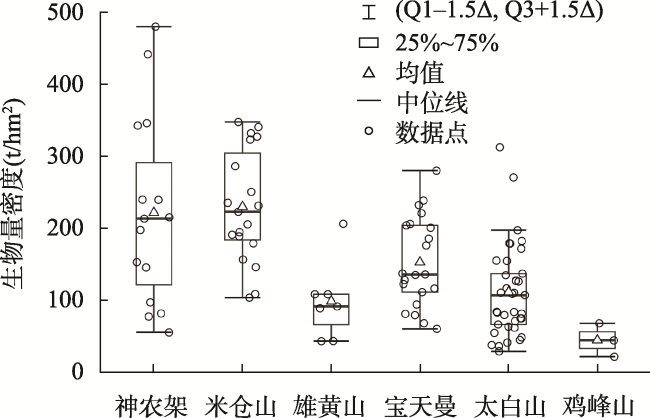
表3 阔叶林样地地上生物量统计Tab. 3 Statistics of aboveground biomass of broadleaved forest based on survey samples (t/hm2) |
| 样地位置 | 样地数量 | 生物量范围 | 生物量均值 | 生物量中位数 | 生物量标准差 |
|---|---|---|---|---|---|
| 东线 | 39 | 10.62~30.1.91 | 79.00 | 53.14 | 62.80 |
| 中线 | 25 | 18.05~249.70 | 139.90 | 126.35 | 63.85 |
| 西线 | 23 | 21.61~275.26 | 109.92 | 84.87 | 75.68 |
| 东西线 | 39 | 9.60~178.82 | 76.64 | 75.63 | 36.03 |
| 佛坪-长青 | 42 | 19.59~416.56 | 162.52 | 139.33 | 111.38 |
| 太白山 | 37 | 28.94~311.88 | 111.05 | 106.64 | 62.79 |
| 米仓山 | 19 | 103.26~346.95 | 229.50 | 204.90 | 75.69 |
| 宝天曼 | 21 | 59.86~279.43 | 152.53 | 135.34 | 61.26 |
| 神农架 | 15 | 54.98~479.21 | 221.21 | 197.08 | 126.54 |
| 雄黄山 | 7 | 42.95~205.68 | 98.13 | 88.39 | 50.66 |
表4 秦巴山区阔叶林地上生物量环境影响因素的相关分析及地理探测器分析结果Tab. 4 Geodetector and correlation analysis results of the environmental factors influencing the broadleaved forest aboveground biomass |
| 气温 | 降水 | 地形 | HAI | |
|---|---|---|---|---|
| q 值(解释力) | 0.191 | 0.079 | 0.163 | 0.120 |
| p 值(0.05显著水平) | 0.000 | 0.014 | 0.000 | 0.000 |
| 皮尔逊相关性 | -0.261** | 0.355** | 0.316** | -0.257** |
| 皮尔逊相关性(>1500 m样本) | 0.226* | 0.476** | -0.191* | 0.152 |
| 显著性(双尾) | 0.000 | 0.000 | 0.000 | 0.000 |
注:*和**分别表示在0.05和0.01级别(双尾)相关性显著。 |
真诚感谢二位匿名评审专家在论文评审中所付出的时间和精力,评审专家对本文研究思路、结果分析、结论梳理方面的修改意见,使本文获益匪浅。也感谢崔璐璐和刘玉莲同学在生物量数据处理方面的工作。
| [1] |
IPCC. Climate change 2021: The physical science basis. https://www.ipcc.ch/report/ar6/wg1/, 2023-12-04.
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
杨元合, 石岳, 孙文娟, 等. 中国及全球陆地生态系统碳源汇特征及其对碳中和的贡献. 中国科学: 生命科学, 2022, 52(4): 534-574.
[
|
| [7] |
|
| [8] |
|
| [9] |
徐新良, 曹明奎, 李克让. 中国森林生态系统植被碳储量时空动态变化研究. 地理科学进展, 2007, 26(6): 1-10.
[
|
| [10] |
周国逸, 尹光彩, 唐旭利, 等. 中国森林生态系统碳储量:生物量方程. 北京: 龙门书局, 2018.
[
|
| [11] |
程堂仁. 甘肃小陇山森林生物量及碳储量研究. 北京: 北京林业大学博士学学位论文, 2007: 18.
[
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
Mohd Zaki Nurul Ain, Abd Latif Zulkiflee. Carbon sinks and tropical forest biomass estimation: A review on role of remote sensing in aboveground-biomass modelling. Geocarto International, 2016, 32(7): 701-716. DOI: 10.1080/10106049.2016.1178814.
|
| [19] |
|
| [20] |
肖瑜. 秦岭地区油松人工林的生物量和产量. 陕西林业科技, 1989, (1): 14-18.
[
|
| [21] |
李晶晶, 党坤良, 周勇, 等. 秦岭中段南坡华山松林能量结构特征和碳储量研究. 西北农林科技大学学报(自然科学版), 2013, 41(11): 58-66.
[
|
| [22] |
傅志军. 太白山太白红杉林的群落学特征及生物量的研究. 汉中师院学报(自然科学版), 1994, (2): 69-72.
[
|
| [23] |
傅志军. 太白山红桦林的群落学特征及生物量研究. 延安大学学报(自然科学版), 1994, 13(2): 37-40.
[
|
| [24] |
李亮, 何晓军, 胡理乐, 等. 1958—2008年太白山太白红杉林碳循环模拟. 生态学报, 2013, 33(9): 2845-2855.
[
|
| [25] |
杨凤萍, 胡兆永, 张硕新. 不同海拔油松和华山松林乔木层生物量与蓄积量的动态变化. 西北农林科技大学学报(自然科学版), 2014, 42(3): 68-76.
[
|
| [26] |
陈存根, 龚立群, 彭鸿, 等. 秦岭锐齿栎林的生物量和生产力. 西北林学院学报, 1996, 11(S1): 103-114.
[
|
| [27] |
程堂仁, 马钦彦, 冯仲科, 等. 甘肃小陇山森林生物量研究. 北京林业大学学报, 2007, 29(1): 31-36.
[
|
| [28] |
王向雨, 胡东, 贺金生. 神农架地区米心水青冈林和锐齿槲栎林生物量的研究. 首都师范大学学报(自然科学版), 2007, 28(2): 62-67.
[
|
| [29] |
尉文. 尺度效应和生境异质性对太白山锐齿栎林物种多样性和地上生物量的影响. 杨凌: 西北农林科技大学硕士学位论文, 2020: 32.
[
|
| [30] |
张九东. 陕西省秦巴山区红豆杉生物量研究. 西安文理学院学报(自然科学版), 2014, 17(2): 19-22.
[
|
| [31] |
李鹏. 基于地统计学的秦岭南坡森林乔木层地上生物量空间分布研究. 杨凌: 西北农林科技大学硕士学位论文, 2013: 19.
[
|
| [32] |
杜宝刚. 宝鸡市森林活立木碳储量、碳密度分布特征研究. 西安: 西北大学硕士学位论文, 2017: 26.
[
|
| [33] |
王耀武. 秦岭(陕西段)生态资产时空变化特征及其驱动因素. 西安: 长安大学硕士学位论文, 2018: 13.
[
|
| [34] |
李梦莹, 胡勇, 曹广强. 基于HJ-1A/B CCD影像的神农架巴山冷杉森林地上生物量估算. 测绘与空间地理信息, 2019, 42(2): 126-130.
[
|
| [35] |
菅永峰, 韩泽民, 黄光体, 等. 基于高分辨率遥感影像的北亚热带森林生物量反演. 生态学报, 2021, 41(6): 2161-2169.
[
|
| [36] |
张百平. 中国南北过渡带研究的十大科学问题. 地理科学进展, 2019, 38(3): 305-311.
[
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548.
[
|
| [42] |
|
| [43] |
王振洲. 离群点检测方法研究及其在机器学习中的应用. 北京: 中国地质大学硕士学位论文, 2018: 30.
[
|
| [44] |
|
| [45] |
|
| [46] |
王劲峰, 徐成东. 地理探测器:原理与展望. 地理学报, 2017, 72(1): 116-134.
[
|
| [47] |
|
| [48] |
徐伟义, 金晓斌, 杨绪红, 等. 中国森林植被生物量空间网格化估计. 自然资源学报, 2018, 33(10): 1725-1741.
[
|
| [49] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}